When you access EPFL websites, we may set cookies on your devices and process personal data about you in accordance with our privacy policy. You can block cookies by using your browser settings.
We are interested in understanding fundamental cellular processes, including in the context of development. Our main focus is on the mechanisms governing asymmetric division and centriole assembly.
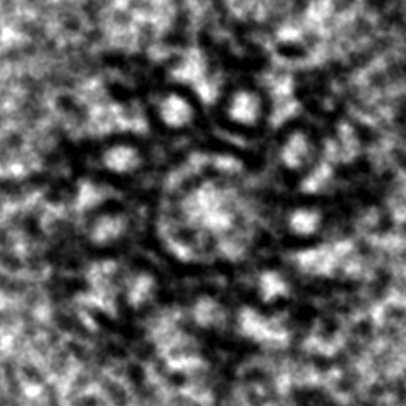
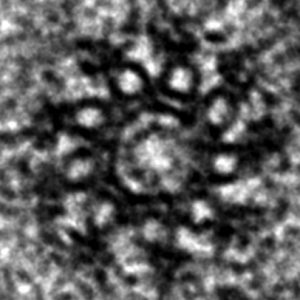
Electron-microscopy of C. elegans centriole during oogenesis viewed in cross-section. Note 9-fold radial symmetric arrangement of microtubule singlets towards the periphery, as well as additional densities, including peripheral-most paddlewheels. The cross-sectional diameter is ~150 nm.2/6
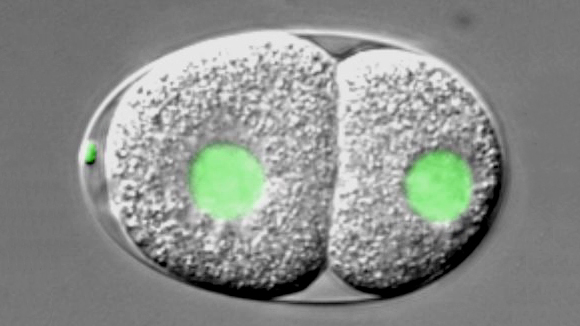
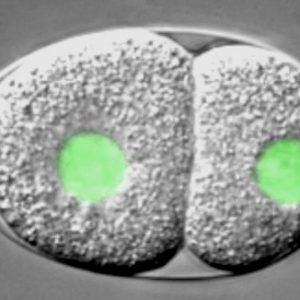
C. elegans embryo carrying a GFP-histone2B fusion protein imaged using dual time-lapse differential interference contrast (DIC) and overlaid fluorescence microscopy,. The embryo is ~50 μm long.3/6
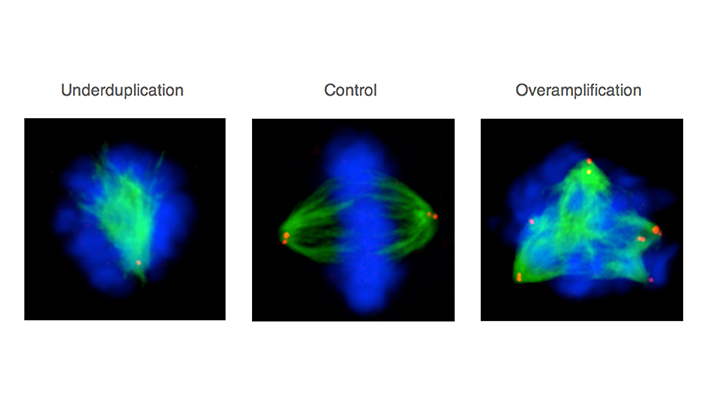
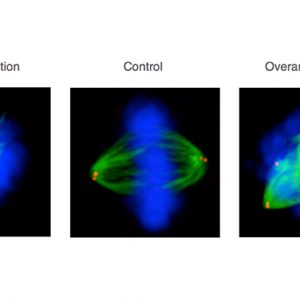
Mitotic HeLa human tissue culture cells stained with antibodies against -tubulin (viewed in green), the centriolar protein centrin (viewed in red), and counterstained with a DNA dye (viewed in blue). Middle: control condition; left: cell depleted by RNAi of a component essential for centriole assembly –note monopolar spindle; right: cell depleted by RNAi of a component essential for restricting centriole assembly –note multipolar spindle. See Balestra et al., 2013. 4/6
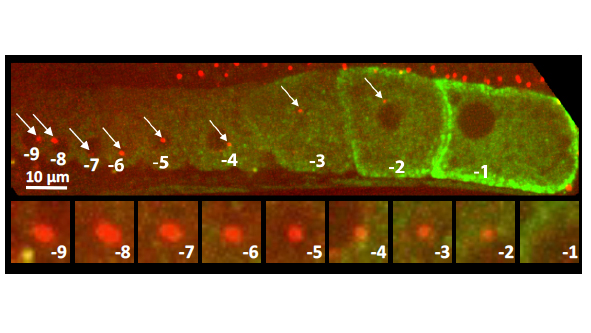
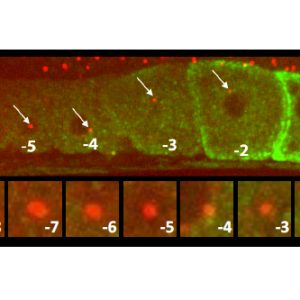
C. elegans gonad from an animal expressing the centriolar marker RFP:SAS-7 and a the marker of oocyte maturation RME-2::GFP; lower row shows inset of centriolar region. Note that the focus of RFP:SAS-7 diminishes as the oocyte matures and is entirely absent from the -1 oocyte.5/6
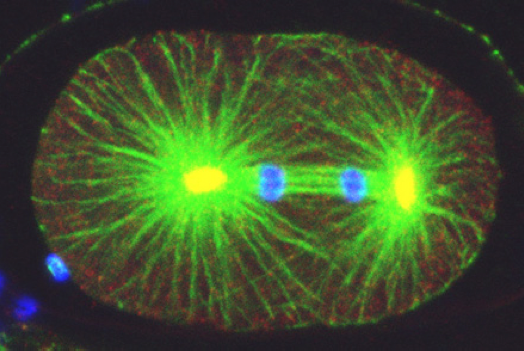
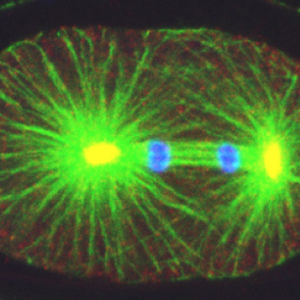
, the centrosomal component ZYG-9 (red, yellow in the overlay with green), and counterstained with a DNA dye (blue). Note asymmetric spindle positioning. © EPFL - Gönczy Lab")