Background:
Over the past decade, there has been a significant emphasis on exploring motor encoding within neural populations. Specifically, research has illuminated the existence of neural activity occurring within relatively low-dimensional subspaces known as neural manifolds. These neural manifolds contain the neural trajectories or patterns associated with various movements or phases of movements. For instance, studies conducted in monkeys have revealed that motor preparation activity, which also activates the motor cortex, and motor generation activity exist within subspaces being orthogonal to each other. Furthermore, the structure of these neural manifolds remains consistent across different tasks. It is only through the analysis of latent activity within these manifolds that one can discern components that are task-dependent as well as those that are task-independent.
Over the past decade, there has been a significant emphasis on exploring motor encoding within neural populations. Specifically, research has illuminated the existence of neural activity occurring within relatively low-dimensional subspaces known as neural manifolds. These neural manifolds contain the neural trajectories or patterns associated with various movements or phases of movements. For instance, studies conducted in monkeys have revealed that motor preparation activity, which also activates the motor cortex, and motor generation activity exist within subspaces being orthogonal to each other. Furthermore, the structure of these neural manifolds remains consistent across different tasks. It is only through the analysis of latent activity within these manifolds that one can discern components that are task-dependent as well as those that are task-independent.
Our focus:
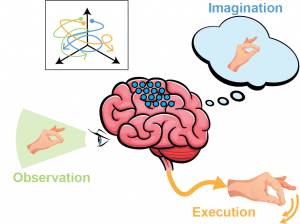
Imagined and observed movements are also known to activate some regions usually associated to motor execution. However, these two conditions do not produce any movement. The question of how this activity is spanned within a lower dimensional manifold hence arises. Are these activities also lying on orthogonal surfaces? Are imagination and observations somehow contained in the more complex manifold spanning execution-related neural activity? Or are these three conditions somehow overlapping on some dimensions, but diverging on others? Can we differentiate the neural trajectories corresponding to different movements within these neural subspace identified?
Imagined and observed movements are also known to activate some regions usually associated to motor execution. However, these two conditions do not produce any movement. The question of how this activity is spanned within a lower dimensional manifold hence arises. Are these activities also lying on orthogonal surfaces? Are imagination and observations somehow contained in the more complex manifold spanning execution-related neural activity? Or are these three conditions somehow overlapping on some dimensions, but diverging on others? Can we differentiate the neural trajectories corresponding to different movements within these neural subspace identified?
Why is this interesting?
– this would shed light on how complex motor behavior is encoded in the cortex. Can we reproduce some of the results found in other species and/or with multi-units activity using Local Field Potential (LFPs) recordings in humans?
– patients having some residual movements could benefit from the development of innovative brain-machine interface taking the interplay between execution and imagination into account
– similarly, innovative brain-machine interface could leverage these findings for the control of new degrees of freedom, like for the control of supernumerary limbs!
– this would shed light on how complex motor behavior is encoded in the cortex. Can we reproduce some of the results found in other species and/or with multi-units activity using Local Field Potential (LFPs) recordings in humans?
– patients having some residual movements could benefit from the development of innovative brain-machine interface taking the interplay between execution and imagination into account
– similarly, innovative brain-machine interface could leverage these findings for the control of new degrees of freedom, like for the control of supernumerary limbs!
References:
- S. Saxena and J. P. Cunningham, “Towards the neural population doctrine,” Current Opinion in Neurobiology, 2019
- J. A. Gallego et al., “Neural Manifolds for the Control of Movement,” Neuron, 2017
- J. P. Cunningham and B. M. Yu, “Dimensionality reduction for large-scale neural recordings,” Nat Neuroscience, 2014
- M. T. Kaufman et al., “Cortical activity in the null space: permitting preparation without movement,” Nat Neuroscience, 2014
- N. Natraj et al., “Compartmentalized dynamics within a common multi-area mesoscale manifold represent a repertoire of human hand movements,” Neuron, 2022
- B. M. Dekleva et al., “Motor cortex retains and reorients neural dynamics during motor imagery”, bioRxiv, 2023.
- G. Dominijanni et al., “Enhancing human bodies with extra robotic arms and fingers: The Neural Resource Allocation Problem”, Nature Machine Intelligence, 2021
- X. Jiang et al., “Structure in Neural Activity during Observed and Executed Movements Is Shared at the Neural Population Level, Not in Single Neurons”, Cell Reports, 2020
Contact
If you are interested in this project and wish to collaborate, please contact me directly
Leonardo Pollina (leonardo.pollina@epfl.ch)